Botanika

A növénytan vagy botanika a biológiának növényekkel foglalkozó ága. A botanika szó ógörög eredetű, a βοτάνη (botané) szó legelőt, füvet, takarmányt jelent.[1][2][3] A botanikusok érdeklődési körébe mintegy 400 ezer faj tartozik,[4] amelyből 248 ezer virágos növény.[5]
A botanika mint a mérgező és gyógynövények ismerete a legrégebbi tudományágak közé tartozik. Az ókortól kezdve egészen a reneszánszig az orvostudomány egyik mellékágaként oktatták; ebből a célból számos füvészkertet is létesítettek az egyetemeken. A növényfajok tudományos rendszerezésének, katalogizálásának igénye már korán felmerült, a jelenleg is használt kéttagú tudományos elnevezéseket Carl Linné vezette be a 18. században.
A 19. és 20. században jelentős fejlődésen esett át a növényi sejtbiológia, genetika és biokémia. A 20. század végén a molekuláris biológia módszereinek felhasználásával lehetővé vált a növények rendszerezése genetikai kódjuk alapján.
A modern növénytan a kutatások igen széles skáláját foglalja magába, többek között tanulmányozza a növények morfológiáját, élettanát, biokémiáját, egyedfejlődését, ökológiáját, evolúcióját, rendszerezését és a növények betegségeit. A botanikai kutatások gyakorlati alkalmazásából a mezőgazdaság, erdészet, textilipar, gyógyszeripar, építőipar stb. profitálhat; segíthetnek megőrizni a biodiverzitást és enyhíteni a globális felmelegedés hatásait.
Története

A botanika mint tudomány gyökerei a gyógynövényismerethez nyúlnak vissza.[6] A legkorábbi növénytani leírások és osztályozások i. e. 1100 előtti indiai,[7] valamint avesztai nyelven írt iráni[8] és olyan kínai szövegekből származnak, amelyeket az ország i. e. 221-beli egyesülése előtt írtak.[7][9]
A modern botanika az ókori Görögországban vette kezdetét, elsősorban Arisztotelész tanítványa, Theophrasztosz (kb. i. e. 371–287) által, aki kidolgozta és leírta alapvető elveit és emiatt „a botanika atyjának” is szokták nevezni.[10] Ebben a témában írt művei, A növények természetrajza és A növények okairól, 1700 évig, egészen a középkorig, a botanika tudományának alapművei voltak..[10][11] Rajta kívül nagy befolyást gyakorolt az időszámításunk első évszázadában élt orvos és gyógyszerész, Pedaniosz Dioszkoridész ötkötetes, gyógynövényekről írt enciklopédiája, a De Materia Medica, amelyet szintén legalább 1500 évig forgattak rendszeresen.[12] A kora középkori iszlám tudomány hozzájárulása a botanikához többek között Ibn Vahsijja Nabateus mezőgazdaság, Abu Hanifa ad-Dinavari (828–896) Növények könyve és Ibn Baszal A talajok osztályozása c. művei. A 13. században Abu al-Abbász al-Nabati és Ibn al-Bajtar (†1248) írt tudományos jellegű növénytani munkákat.[13][14][15]
A 16. század közepén több itáliai egyetemen botanikus kertet létesítettek; az elsőt a hagyomány szerint Padovában, amely ma is 1545-ben kialakított helyén található. Korábban a kolostorok tartottak fenn praktikus okokból gyógynövénykerteket. Az egyetemi botanikus kertek segítették a tudományág oktatását és gyógyító hatásuk demonstrálását. Észak-Európában csak később terjedtek el, pl. Angliában az első botanikus kertet az Oxfordi Egyetemen hozták létre, 1621-ben. Ekkoriban a botanika még mindig csak az orvostudomány mellékága volt.[16] Németországban Leonhart Fuchs (1501–1566) és Hieronymus Bock orvosok (1498–1554) (utóbbit Hieronymus Tragusként is említik), valamint Otto Brunfels teológus (1489–1534) volt a botanika három német atyja[17] Fuchs és Brunfels szakított a hagyománnyal, amely az ókori írók másolásán alapult és saját megfigyeléseket is tettek; Bock pedig kidolgozta saját osztályozási rendszerét. A szintén német Valerius Cordus (1515–1544) halála évében adta ki a botanikai és gyógyszerészeti szempontból egyaránt jelentős Historia Plantarumot, két évvel halála utána pedig megjelent a sokáig használatban lévő gyógyszerkönyve, a Dispensatorium.[18] A kor jelentős botanikusai voltak a svájci Conrad von Gesner (1516–1565), az angol John Gerard (1545–1611) és az olasz Ulisse Aldrovandi (1522–1605). Robert Hooke 1665-ben mikroszkópjával a parafát vizsgálva fedezte fel a sejteket, amelyeket később élő növényi szövetekben is felismert.[19]
Az első magyar füveskönyvet Méliusz Juhász Péter (1532-1572) debreceni református lelkész adta ki Herbarium az füveknek nevekről, természetekről és hasznairól címmel. Könyve Adam Lonitzer Herbariumán alapult, és bár kiegészítette magyarországi növényekkel is, ő maga nem volt botanikus. Beythe István (1532–1612) 1583-ban összeírta a növények magyar és latin neveit és Stirpium nomenclator Pannonicus Németújváron adta ki. Fia, a szintén református pap Beythe András (1564-1599) Melius könyvére alapozva szintén írt egy népszerű füveskönyvet.[20] Lásd még: A magyar botanika története

A 18. században Európában kezdték megismerni a távoli kontinensek, gyarmatok növényeit is. Megjelentek az első taxonómiai rendszerek (rend, család stb.) és elkészültek az első választásos kulcs szerinti növényhatározók is. A korai rendszerek sok esetben mesterséges csoportokba osztották be a növényeket. 1753-ban Carl von Linné Species Plantarum címmel megjelentette saját, hierarchikus osztályozási rendszerét, amely máig a növénytaxonómia alapját képezi. Ő vezette be a kéttagú latin tudományos neveket, amelyben az első név a nemzetséget, a második a fajt jelentette.[21] Linné a porzók száma alapján rendszerezte 24 csoportba a virágos növényeket, és valamennyi virág és porzó nélküli fajt (mohákat, páfrányokat, gombákat) a Cryptogamia csoportba sorolta.[22]
A növényanatómiai, morfológiai és az életciklusokra vonatkozó ismeretek gyarapodásával továbbfejlesztették Linné mesterséges rendszerét. Adanson (1763), De Jussieu (1789) és De Candolle (1819) a virágzaton túl további jellegzetességekkel jellemezte a taxonómiai csoportokat. Ekkoriban az osztályozást pusztán morfológiai alapon végezték, és kényelmi, gyakorlati szempontok vezérelték; miután azonban elterjedt Darwin evolúcióelmélete, a rokon csoportokat egymás tényleges leszármazottainak kezdték tekinteni.[23]
Az első modern botanikai mű Matthias Schleiden 1842-ben kiadott Grundzüge der Wissenschaftlichen Botanikja volt.[24] Schleiden sokat mikroszkopizált és Theodor Schwann-nal valamint Rudolf Virchowval közösen kidolgozta az élőlények sejtelméletét; továbbá az elsők között ismerte fel a Robert Brown által felfedezett sejtmag jelentőségét.[25]
Modern növénytan

A 19. század közepén Gregor Mendel (1822-1884) borsóval végzett kísérletekkel bizonyította be a gének létezését. A század végén Eugenius Warming és más tudósok munkája révén kialakultak a növényökológia alapkoncepciói, rájöttek, hogy a növények társulásokat alkotnak. Christen C. Raunkiær elmélete a növények életformáiról a mai napig érvényben van. Henry Chandler Cowles, Arthur Tansley és Frederic Clements rájött, hogy a társulások idővel megváltozhatnak, vagyis felfedezték az ökológiai szukcesszió jelenségét. Clements vezette be a klimax vegetáció fogalmát, amely az adott klimatikus viszonyoknak leginkább megfelelő, legstabilabb társulást jelenti. Alphonse de Candolle korábbi munkáira alapozva, a szovjet Nyikolaj Ivanovics Vavilov lefektette a biogeográfia alapjait, bevezette a haszonnövények eredetcentrumainak fogalmát, és felderítette háziasításuk folyamatának részleteit.[26] A 20. század elején Katherine Esau (1898–1997) adta ki növényanatómiai könyveit, amelyek majdnem egy félszázadig voltak nélkülözhetetlen alapművek.[27][28]
Az 1960-as évek közepétől fény derült a növényélettan fizikájára, ismertté vált a vízszállítás mechanizmusa, a párologtatás hőmérsékletfüggése és a sztómákon keresztül történő vízpára- és szén-dioxid-diffúzó mikéntje.[29][30] A Ronald Fisher,[31] Frank Yates és más kutatók által bevezetett statisztikai újítások a botanikában is átalakították a kísérlettervezést és a kapott adatok analízisét.[32] 1948-ban Kenneth V. Thimann felfedezte az első növényi hormont, az auxint, amellyel be lehetett avatkozni a növények fejlődésébe. Frederick Campion Steward kifejlesztette a hormonok által szabályozott növényi szövetkultúra módszerét.[33] A szintetikus auxin (2,4-dikloro-fenoxiecetsav) egyike volt az első kereskedelmi forgalomba kerülő szintetikus herbicideknek.[34]
A szerves kémiai analízis 20. századi fejlődése, az olyan technikák, mint a spektroszkópia, kromatográfia vagy elektroforézis nagyban elősegítették a növényélettan biokémiájának kutatását. Tanulmányozhatóak a növények génjei, az azokról készülő fehérjék, valamint anyagcseréjük részletei.[35] Gottlieb Haberlandt már 1902-ben feltételezte,[36] hogy valamennyi növényi sejt totipotens, vagyis szövetkultúrában egész növénnyé fejleszthető. A jelenség kihasználásával egyszerűen lehet tanulmányozni a gének hatásait, azáltal hogy kikapcsolják őket vagy jelzőmolekulákat (mint pl. a zöld fluoreszcens protein) kötnek hozzájuk. A modern génsebészeti technikákkal előállított növények vagy szövettenyészetek képesek peszticideket, antibiotikumokat vagy egyéb hatóanyagokat termelni.[37] A taxonómia is egyre inkább felhagy a morfológiai tulajdonságok elemzésével és helyette a DNS-szekvencia alapján határozzák meg a fajok rokonsági viszonyait. A nukleinsav-analízis alapján 1998-ban újragondolták a zárvatermők családjainak egymás közötti rokonságát.[38]
A növénytan jelentősége és ágazatai

Mivel a növények termelik szinte a teljes állati élet létfenntartásához szükséges oxigént és kémiai energiát, tanulmányozásuk alapvető fontosságú. A soksejtű növényekben és egysejtű algákban (valamint a baktériumokhoz tartozó kékmoszatokban zajlik a fotoszintézis folyamata, amely a napfény energiája, szén-dioxid és víz felhasználásával energiahordozó és a szintetikus folyamatokban felhasználható cukrokat állít elő.[39][40] A fotoszintézis melléktermékeként oxigén szabadul fel, amelyet (egyes baktériumok kivételével) valamennyi élőlény felhasznál a létfolyamataihoz szükséges kémiai energia előállításához. A növények fontos szerepet játszanak a szén és a víz globális körforgásában, gyökereik megkötik és stabilizálják a talajt, megakadályozva azok erózióját.[41] A növények alapvető fontosságúak az emberiség fennmaradása és jövője szempontjából: élelem, oxigén, gyógyszerek, építőanyag, ruhák, papír és számos más iparcikk forrásai.[42]
A botanikusok a növények biokémiai folyamatait, sejtműködésüket, szöveti felépítésüket, az egyes egyedeket vagy társulásaikat tanulmányozzák. A tudományághoz tartozik a növények osztályozása és törzsfejlődésének tanulmányozása is.[43]
Korábban az élővilágot állatokra és növényekre osztották fel[44] és a növénytanhoz soroltak mindent, ami nem volt állatnak minősíthető, vagyis a gombákat és baktériumokat is.[45] A szó legszigorúbb értelmezésében a „növény” a szárazföldi növényeket vagy embriofitákat jelenti, amely a zárvatermőket (pl. virágos növények), nyitvatermőket (fenyők), harasztokat (páfrányok) és mohákat foglalja magába. Az embriofiták soksejtű algáktól származnak és életciklusokban kisebb-nagyobb mértékben a mai napig megfigyelhető a moszatokban kialakult haploid (gametofiton) és diploid (sporofiton) generációk váltakozása. A fejlettebb növényeknél (nyitva- és zárvatermők) a gametofiton már csak minimális méretű és a sporofitontól csak mikroszkóp alatt lehet elkülöníteni.[46] A baktériumokat ma a mikrobiológia, a gombákat és zuzmókat a mikológia, a nem-zöld algákat a fikológia, a vírusokat a virológia tanulmányozza. Ettől függetlenül a botanikusnak készülő egyetemi hallgatókat többnyire megismertetik a gombákkal és fotoszintetizáló egysejtűekkel is.[47][48]
Az ősnövénytan (paleobotanika) a kihalt növényeket tanulmányozza kövületeik alapján, hogy információkat nyerjen a növényvilág törzsfejlődéséről. Mai tudásunk szerint a növények ősi eukarióta sejtek és fotoszintetizáló kékmoszatok endoszimbiózisa révén jöttek létre, amikor utóbbiakból kloroplasztiszok lettek.
Az etnobotanika a néprajz és a növénytan határán elhelyezkedő tudományág, kutatási területe a növényfajok emberi kultúrákra gyakorolt hatása ma és a múltban. [49]
A botanika olyan 21. századi feladatokhoz nyújthat segítséget, mint a környezetvédelem, az élelemellátás biztonsága, az invazív fajok kontrollálása, a klímavédelem (és ezzel kapcsolatban a szén lekötése a légkörből).[50]
Élelmiszerellátás

Valamennyi élelmünk vagy közvetlenül a növényekből, vagy közvetetten, az őket elfogyasztó állatokból származik.[51] A fotoszintézis miatt a növények képezik szinte minden táplálkozási láncolat legelső fokát.[52] A legfontosabb élelmiszernövények a búza, a kukorica, a rizs, a banán, a hüvelyesek, vagy a ruhát szolgáltató gyapot, mind több ezer évvel ezelőtt lettek nemesítve vadon növő elődeikből. [53] A genetika és élettan előrehaladtával lehetővé vált olyan új fajták létrehozása, amelyekkel jóval nagyobb termés érhető el.[54] A botanikusok a gyomnövényeket és növénypatogén organizmusokat is tanulmányozzák, amelyek komoly károkat képesek okozni mind a mezőgazdaságban, mind a természetes ökoszisztémákban.[55]
Növényi biokémia
A növények (és más fotoszintetizáló eukarióta szervezetek) egyedi fotoszintetizáló organellumokkal, kloroplasztiszokkal rendelkeznek. A kloroplasztiszok (és elődeik, a kékalgák) kékeszöld (klorofill-a) és sárgászöld (klorofill-b) pigmentet tartalmaznak[56]. A kékalgákban és egyes moszatcsoportokban másfajta klorofillformák is előfordulnak, de a szárazföldi növények ezeket nem tartalmazzák.[57][58][59] Ezek a festékanyagok elnyelik a spektrum kék-ibolya és narancs-vörös részét, míg a zöld fényt visszaverik, ezért zöldek a növények. Az elnyelt kék és vörös fény energiáját felhasználva a kloroplasztiszokban energiadús szénhidrátmolekulák képződnek és a folyamat melléktermékeként oxigén szabadul fel.

A fény energiája kilök a klorofillból egy elektront, amely kémiai átalakulásokat indít el, aminek a végén a kloroplasztisz membránján protongrádiens (a két oldala közötti protonkoncentráció-különbség) alakul ki. Ennek energiájából jön létre az adenozin-trifoszfát molekula, amely a szén-dioxidot a cukrokba fixáló Calvin-ciklust hajtja. A ciklus központi enzime, az ún. RuBisCO, amely ötszénatomos cukrormolekulából egy szén-dioxid beépítésével két három szénatomos gliceraldehid-3-foszfátot képez. Ezek közül az egyik bekerül a glükózszintézisbe, a másik pedig regenerálja a kiindulási ötszénatomos cukrot. A glükóz aztán általánosan használt energiahordozó a sejt életfolyamatiban és keményítő formájában tárolásra is kerül. [60] A legtöbb növény és alga keményítőként tárolja az energiát; ez alól kivételek az őszirózsafélék, amelyek a fruktózpolimer inulint használják erre a célra. A glükóz egy része jól szállítható szacharózzá (közönséges étkezési cukorrá) alakul.
A kloroplasztiszok meglehetős autonómiával rendelkeznek a sejten belül, saját maguk szintetizálják valamennyi számukra szükséges zsírsavat,[61][62] és a legtöbb aminosavat is.[63] A zsírsavakat főleg a lipidmembrán és a kutin polimer készítésére használják fel. [64]
A növények számos, csak rájuk jellemző polimert szintetizálnak, mint pl. az olyan poliszacharidokat, mint a cellulóz, pektin vagy xiloglükán;[65] amelyekből a sejtfal épül fel.[66] Az edényes növények lignint készítenek, amellyel a háncs szállítósejtjeinek a falát szilárdítják meg. A fás növények szintén a lignint alkalmazzák a cellulóz mellett a belső támasztószerkezetük (a faanyag) megerősítésére. A spórák és pollenszemek külső falában található a sporopollenin, amely ellenállóvá teszi a részecskéket a környezeti behatásokkal szemben. Az ordovíciumi kövületekben való megjelenése jelzi a növények szárazföldi térhódítását.[67] A korai földtörténeti korokban a szén-dioxid légköri koncentrációja magasabb volt a mainál. Egyes kétszikű (mint az őszirózsafélék) és több egyszikű növény (kukorica, ananász) egymástól függetlenül szén-dioxidtakarékos módjait fejlesztették ki a szénfixációnak (ún. C4-út, szemben a hagyományos C3-úttal).
Hatóanyagok és ipari alapanyagok

A növények által termelt másodlagos, nem létfontosságú anyagokat a fitokémia tanulmányozza.[68] Ezen vegyületek némelyike mérgező, mint például a bürök és a sárga kürtvirág által termelt koniin alkaloid. Mások az illatszerként felhasznált illóolajuk, vagy fűszerként való alkalmazásuk (pl. kapszaicin) miatt hasznosak, esetleg a gyógyászatban használhatóak, mint a mák ópiumának sok összetevőjét. A hatóanyagok egy részét legális vagy illegális drogként fogyasztják, mint a marihuána tetrahidrokannabinolját, a koffeint, a morfint vagy a nikotint. Néha a tényleges hatóanyag egy növényi termék kis mértékben módosított változta, mint az acetilszalicilsav, amely a fűzfák kérgében megtalálható szalicilsav acetilésztere. [69] Bár nem közvetlenül, de az alkoholos italok is a növények (szőlő, árpa, rizs) által termelt cukrok etanolos erjesztéséből származnak.[70]
A szintetikus festékanyagok felfedezéséig a festékek nagy része növényi eredetű volt. Sárga festéket nyertek a festő rezedából, vöröset a festő buzérból, kéket a festő indigóból vagy festő csüllengből.
Növényi termékek az olyan ipari nyersanyagok, mint a keményítő, a gyapot, len, kender, fa és papír, különböző olajok és viaszok és a természetes gumi. A fából pirolízissel nyert faszenet évezredek óta használják a kohászatban, szűrőanyagként vagy valamivel később mint a lőpor egyik alapanyagát. A világ legelterjedtebb szerves polimerje a cellulóz,[71] hőenergiaként, üzemanyagként, építőanyagként vagy vegyipari alapanyagként hasznosított. Cellulózból készül a műselyem, a celofán, a tapétaragasztó vagy a lőgyapot. Cellulózból, repceolajból, kukoricakeményítőből vagy nádcukorból ipari módszerekkel bioüzenanyag is nyerhető.[72]
Növényökológia

A növényökológia a növények és (élő vagy élettelen) környezetük kölcsönös kapcsolatát tanulmányozza. A növényökológusok a helyi és regionális flóra összetételét tanulmányozzák, a növénytársulásokat, azok faji és a fajokon belül a genetikai diverzitást, a növények alkalmazkodását a környezethez és a fajok egymással való versengését vagy együttműködését.[73] A tudományág célja, hogy megértse a növények eloszlását, produktivitását, környezeti hatását és a környezeti változásokra adott válaszát.[74]
A legfontosabb környezeti tényezők a talaj és az éghajlat (hőmérséklet és csapadék) milyensége. A növények azonban maguk is megváltoztathatják ezeket a faktorokat, például csökkentik a talaj fényvisszaverő-képességét, megkötik a lehulló csapadékot, stabilizálják az ásványi talajokat és növelik azok szervesanyag-tartalmát. A társulásokon belül a növények versengenek az energiaforrásokért és tápanyagokért.[75][76] Az adott környezeti viszonyokhoz leginkább alkalmazkodni képes növények (és állatok) társulásai hozzák létre a jellegzetes biomokat, mint például a tajgát vagy a trópusi esőerdőt.[77]

A növények lehetőségeikhez képest igyekeznek védekezni az ellen, hogy az állatok megegyék őket. Egyes növények parazita életmódhoz alkalmazkodtak, vagy esetleg húsevők. Más fajok kölcsönösen előnyös kapcsolatot alakítottak ki gombákkal (gyökérkapcsoltság, mikorrhiza) vagy állatokkal; főleg hangyákkal, amelyek megvédik az őket tápláló növényeket a parazitáktól, [78] de ide sorolhatóak a virágok beporzását végző rovarok vagy denevérek [79][80] vagy a magok szétszórásában segédkező gyümölcsevő állatok is.[81]
A növényfajok hőmérséklet- és vízigényének ismeretében a maradványaik (különösen az ellenálló pollen) alapján következtetni lehet az egyes régiókban korábban uralkodó klimatikus viszonyokra. A pollenanalízis akár százmillió évere visszamenőleg is adhat támpontot az akkori éghajlatra vonatkozóan.[82] A régi földtörténeti korok légkörének szén-dioxidtartalmára is a levélfosszíliák pórussűrűsége, valamint alakja és mérete alapján következtetnek.[83] Megfigyelték, hogy a magaslégköri ózon csökkenése és az UV-B sugárzás megerősödése gátló hatással van a növények növekedésére.[84] A társulások változásának ismeretével információkhoz juthatunk a klíma változásával vagy az élőhely pusztulásával kapcsolatban.[85]
Genetika
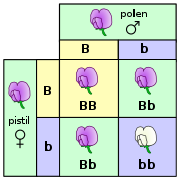
A növények tulajdonságainak öröklődése alapvetően ugyanazokat az elveket követi, mint amik a többi soksejtes élőlényben érvényesek. A 19. században Gregor Mendel a borsót tanulmányozva fedezte fel a géneket és az öröklődés általános törvényeit. A 20. században Barbara McClintock a kukoricában ismerte fel az ugráló gének, a transzpozonok létezését. [86] Vannak azonban olyan genetikai tulajdonságok, amelyek a növényekre jellemzőek.
A fajközi genetikai elkülönülés jelentősen gyengébb, mint az állatok esetében és gyakoribbak a fajok közötti hibridek. A jól ismert borsmenta például a vízi menta (Mentha aquatica) és a fodormenta (Mentha spicata) steril hibridje.[87] A búza több vad búzafaj bonyolult hibridizációjának eredménye: a nemesített búzában három faj genetikai állománya található meg.[88]
A kétivarú virágokat hozó zárvatermők sok esetben különböző mechanizmusokkal megakadályozzák az önbeporzást, vagyis, hogy a saját pollenjük elérje a bibét vagy létrejöjjön a megtermékenyítés.[89] Az egyszerűbb módszer, amit szintén sok faj követ, hogy a hím- és nőivarú virágok különböző egyedeken nőnek.
A fejlettebb állatoktól eltérően a növények esetében igen gyakori a vegetatív, aszexuális szaporodás. Ilyen a burgonya gumóin lévő sarjrügyek vagy a földieper legyökerező indái általi szaporodás. Különösen elterjedt ez a sarkvidékei vagy magashegyi környezetben, ahol a rovarbeporzás lehetősége korlátozott és a növények virágok helyett sokszor sarjhagymákat vagy -rügyeket hoznak.
A legtöbb szexuális úton szaporodó élőlény diploid, vagyis minden kromoszómából két példánnyal rendelkezik. Néha, a sejtosztódás folyamatának hibája folytán a kromoszómaszám megtöbbszöröződik, vagyis a sejt poliploid lesz. Ha ez az egyedfejlődés korai szakaszában történik, a szervezet teljesen vagy részlegesen autopoliploid lesz, vagy hibridizáló ivarsejtek esetén allopoliploid. Utóbbi esetben az utód nagyobb eséllyel lesz életképes, mert az elkövetkező sejtosztódások során minden kromoszómájának lesz párja. Az állatok esetében a poliploidok szinte soha nem életképesek, ám a növények sokkal jobban viselik a többszörös kromoszómaszámot. Mivel ezek az egyedek az eredeti, diploid társaikkal nem tudnak szaporodóképes utódot létrehozni, többnyire vegetatívan szaporodnak és ha sikeresnek bizonyulnak, általuk új faj jöhet létre. [90] A kereskedelemben kapható banán ilyen steril, magot nem termelő triploid hibrid. A pitypangnak Észak-Amerikába behurcolt változata szintén triploid, amely a magvak apomixise révén produkál csírázó magvakat.
A kloroplasztiszok és mitokondriumok öröklődése nem mendeli jellegű, a nyitvatermőknél a porzós, míg a virágos növényeknél a termős elődtől származnak.[91]
Molekuláris genetika

A klasszikus és molekuláris genetika egyik modellszervezete a káposztafélék családjába tartozó, kis termetű lúdfű (Arabidopsis thaliana).[92] Ez volt az első növény, amelynek teljes genomját megszekvenálták. [93] Kromoszómáinak DNS-e kb 135 millió bázispár hosszú, ami a virágos növények között a legkisebb genomok közé tartozik. Az eredmények fontos információkat szolgáltattak a növények genetikájáról és molekuláris felépítéséről, akárcsak az azóta elvégzett újabb teljesgenom-szekvenálások a rizs (Oryza sativa)[94] vagy a gabonafélékkel rokon Brachypodium distachyon esetében.[95]
Az ideális modellnövénynek kevés génje van, kicsi, könnyen termeszthető és rövid a generációs ideje. Bár ezeknek a feltételeknek nem mindenben felel meg, gazdasági jelentősége miatt sokat foglalkoztak a kukorica genetikájával és a rá jellemző C4-típusú fotoszintézissel.[96] A fotoszintézis és a kloroplasztiszok kutatásában nagy jelentőségre tett szert a Chlamydomonas reinhardtii egysejtű zöldalga [97] és a vörösmoszatokhoz tartozó Cyanidioschyzon merolae.[98] A növényi sejtműködés kutatásában visszatérő tesztnövények a spenót,[99] a borsó,[100] a szója vagy a Physcomitrella patens mohafaj.[101]
Az Agrobacterium tumefaciens talajbaktérium a gyökerekhez kapcsolódva képes beléjük juttatni egy plazmidot, amely a sejteket burjánzásra ösztönzi és gümőt hoz létre. Már 1977-ben felmerült, hogy a plazmidot fel lehetne használni arra, hogy a nitrogénfixációhoz szükséges géneket bevigyék a növényi sejtekbe. Ma a Ti plazmid a transzgénikus növények létrehozásának fontos eszköze.[102]
Az epigenetika az olyan öröklődési mechanizmusok tudománya, amelyeket nem lehet a hagyományos génmodellel megmagyarázni[103] Ilyen lehet az, amikor a petesejtben a DNS metilálása vagy bizonyos fehérjék kötődése szabályozza a gének kifejeződését. Az ilyen szabályozás okozhatja a levelek, szirmok különbözőségét akkor is amikor a genetikai háttér azonos. Az epigenetikai változás lehet ideiglenes vagy az egyed egész életén át tartó, sőt egyes esetekben akár az utódokra is átörökített[104]; bár az ivarsejtek érésekor a gének szabályozása többnyire visszaállítódik az eredeti állapotba.
Az állatoktól eltérően a legtöbb növényi sejtben a szervezet fejlődése során lezajló epigenetikai változások visszafordíthatóak, vagyis kedvező körülmények között a sejtből az egész növény kinöveszthető. Kivételek lehetnek az elfásodott szállító- vagy támasztósejtek, vagy a floém szitasejtjei, amelyek elveszítik sejtmagjukat.
A növények evolúciója
Ma általánosan elfogadott, hogy a növények kloroplasztiszai valaha autonóm kékmoszatok voltak; erre számos genetikai, biokémiai és morfológiai bizonyítékot találtak. A kékmoszatok a korai eukarióta sejtek endoszimbiótái lettek és idővel teljesen elvesztették önállóságukat.[105][106][107][108]
Az algák egy nagy és változatos polifiletikus csoport. Az egyes alcsoportok között biokémiájukban, pigmentjeikben, sejtfelösszetételükben, kloroplasztiszszerkezetben, tápanyagokban elég nagyok az eltérések. A szárazföldi zöld növények a zöldmoszatok (Chlorophyta) közeli rokonaitól, a csillárkamoszatoktól (Charophyta) származnak.[109] Az utóbbiak egyik osztálya, a Charophyceae és az embriós növények egyetlen, közös eredetű ágazatot alkotnak, a Streptophytinákat.[110]
A legősibb szárazföldi növényekben még nem fejlődtek ki a szállítószövetek, ők az ún. edénytelenek, amikhez a lombos-, becős- és májmohák tartoznak. A szilur korszakban jöttek létre az első edényesek, amelyekben már megfigyelhető a xilém és floém elkülönülése. Ebből a korszakból származnak a ma is létező korpafüvek. A devon korszak végén több csoport (korpafüvek és a már kihalt Sphenophyllales és Progymnospermopsida csoportok) egymástól függetlenül kifejlesztették a korábbinál jóval nagyobb megaspórákat és a megaspórákból kifejlődő gametofitonok a spóratokon belül maradtak. Ebből a képletből fejlődtek ki a magvak. Az első magvas növények a devon végén jelentek meg, [111][112] belőlük jöttek létre a már kipusztult magvaspáfrányok, a zárvatermők és a nyitvatermők.[113] A nyitvatermők (fenyők, gingkó, cikászok) szabadon álló magkezdeményeivel szemben a zárvatermők magházzal védik magvaikat.[114][115] A molekuláris genetikai vizsgálatok tanúsága szerint a mai nyitva- és zárvatermők nem egymásból fejlődtek ki, hanem evolúciójuk egymástól független volt.[116]
Növényélettan

A növényélettan az életfolyamatokat kísérő fizikai és kémiai változásokat vizsgálja.[117] Az anyagcseréhez szükséges vegyi anyagokat a növények a levegőből és a talajból szerzik be. A kellő energiát a napfény szolgáltatja, vagyis a legtöbb növény ún. fotoautotróf. Egyes, teljesen parazita növények más növényektől vagy gombáktól szerzik be a szükséges tápanyagokat, ők az állatokhoz és gombákhoz hasonlóan heterotrófok.[118] A fotoszintézis során azonban nem közvetlenül hasznosítható kémiai energiahordozók jönnek létre, hanem olyan szénhidrátok, amelyeket a növényi sejt az állatokéhoz hasonlóan lebont és ATP-vé alakít át; utóbbit már közvetlenül tudja hasznosítani a biokémiai reakciókban.[119]
A növényi szervezeten belül a molekulák mozgatása különböző fizikai folyamatok felhasználásával történik. A sejtmembránokon keresztül ionok, kisebb és nagyobb molekulák mozognak aktív (energiaigényes) és passzív transzport révén. A víz és az ásványi anyagok a gyökerekből a növény többi részébe a párologtatás által hajtott áramlás miatt kerülnek. A szervezet az anyagtranszportban olyan fizikai-kémiai folyamatokat is felhasznál, mint a diffúzió vagy ozmózis.[120] A növények növekedéséhez, a sejtek felépítéséhez vízen és szén-dioxidon kívül szükség van nitrogénre, foszforra, káliumra, kalciumra, magnéziumra és kénre is. Az edényes növények ezeket az elemeket a talajból szerzik be vízben oldható, ionos formában (ahová a talajásványok lebomlásával jutnak) és a szállítószövetek (xilém) segítségével a szervezet minden részébe eljuttatják.[121] A fotoszintézis során a levelekben keletkező cukrot a floém szállítószövet transzportálja a nem-fotoszintetizáló sejtekbe.

.
A növények aktívan reagálnak a külső ingerekre: a fényirányra, érintésre, sérülésre és képesek az adott inger felé vagy attól elmozogni vagy -nőni. Reakciókészségük leglátványosabb példái a szemérmes mimóza levelei vagy a húsevő növények (vénusz légycsapója, hízóka) rovarfogó mozgása[122]
Növényi hormonok
Már a 19. század végén felvetetették, hogy a növények növekedését kémiai faktorok, növényi hormonok szabályozzák. Darwin is kísérletezett a hajtások fény felé és a gyökerek lefelé való irányulásával.[123] Az 1930-as években a holland Frits Warmolt Went bizonyította be az első növekedési hormon, az auxin létezését, amely a fény felé hajlást szabályozza.[124] Hormonokkal sikerült apró szövetdarabokat sejtosztódásra és gyökerek és hajtások növesztésére bírni: ezek voltak a kezdeti lépések a növények biotechnológiájában és génsebészetükben[125]
A későbbiekben felfedezték a sejtosztódást szabályozó citokinineket (elsőként a kukoricában a zeatint). A zeatin a gyökerekben képződik, a floém felviszi a hajtásokba, ahol elősegíti a sejtosztódást, rügyképződést és a növeli a kloroplasztiszok klorofilltartalmát.[126][127] A gibberellinek (pld gibberellinsav és diterpének) a magképződést és a magok csírázását szabályozzák és termelődésüktől függ a kifejlett növény magassága is.[128] A szinte valamennyi szárazföldi növény által termelt abszcizinsav gátolja a sejtosztódást, elősegíti a magok érését és nyugalmi állapotát és bezárja a levelek sztómáit. [129] Az etiléngáz a gyümölcs érését gyorsítja meg és megindítja a levélhullást.[130][131]
Növényanatómia és morfológia
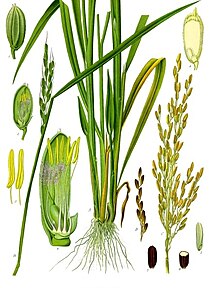
A növényanatómia a szövetek és sejtek belső struktúráját, míg a morfológia külső jellegzetességeiket, alakjukat tanulmányozza.[132] A növényi sejtet alapvetően a cellulózból, hemicellulózból vagy pektinből álló sejtfala különbözteti meg az állati vagy gombasejtektől.[133] Ezenkívül vakuólumai nagyobbak mint az állati sejtekben és plasztidokat tartalmaz. A plasztidok közül a kloroplasztisz a fotoszintézis színhelye míg mások tartalék tápanyagokat, keményítőt (amiloplasztisz) vagy olajt (elaioplasztisz) tartalmaznak. A növényi sejtek osztódása is sajátos, a leánysejteket egy sejtlemez választja el, amely a növekedése során kiszorítja a síkjára merőleges, mikrotubulusokból és az endoplazmatikus retikulum darabjaiból álló fragmoplasztot.[60]

A növények többsége (a mohák kivételével, amelyeknek csak rögzítő rhizoidjuk van) különböző szerkezetű föld fölötti szárral és föld alatti gyökérrel rendelkezik. A száron nőnek a fotoszintézist végző levelek és a reprodukciós szervek. A klorofill nélküli gyökereken apró hajszálgyökerek végzik a víz és ásványi anyagok felszívását.[134] A két szervrendszer egymástól függ: a gyökerek a vizet és ásványi anyagot, a szár és levelek az energiát biztosítják a szervezet számára.[134] Mindkét rendszer képes a másikká átalakulni:[135] a szárhoz tartozó indák és gumók gyökereket ereszthetnek, a sekély mélységben szétterülő gyökerekből (pl. fűz) pedig hajtások nőhetnek ki.[136] Ha valamilyen baleset miatt az egyik rendszer elvész, a másik újranöveszthető; sőt pl. a fokföldi ibolya esetében egyetlen levélből is kinőhet az egész növény[137] Laboratóriumi körülmények között egyetlen sejtből is lehet teljes növényt produkálni[135] A két rendszer között a szállítószövetek tartják a kapcsolatot: a xilém a gyökerekből szállít vizet, a floém a levelekből cukrot. A gyökerek sok esetben tápanyagraktározó funkciót is ellátnak, mint a cukorrépa vagy petrezselyem esetében. [136]
A szár elsődleges funkciója, hogy a levelek és ivarszervek állványzata legyen, de tárolhat vizet (kaktuszok), tápanyagot (burgonya) vagy lehet a vegetatív szaporodás eszköze (földieper).[138] A levelek funkciója a fotoszintézis.[139] A nagy, szétterülő, hajlékony leveleket lomblevélnek; a hosszú, merev, fonalszerűeket pedig tűlevélnek nevezik.[140] A fás növények szöveteiben egy másodlagos differenciáció is lezajlik: a xilémből belülről támasztó, a floémből kívülről védő kéreg lesz. Valamennyi ma élő nyitvatermő fás növény, míg a zárvatermőknek csak egy része.[141]
Növényrendszertan

A rendszertan vagy taxonómia a fajok csoportokba sorolásával foglalkozik, lehetőség szerint evolúciós rokonságuk alapján.[142] A mai rendszertan alapjain Carl Linné fektette le a 18. században, aki külső jegyeik alapján rendszerezte az élőlényeket; ma is az általa bevezetett binomiális nómenklatúra van használatban. Később, Darwin evolúciós elmélete után fontossá váltak a csoportok közti leszármazási viszonyok is. Az utóbbi évtizedekben egyre nagyobb a jelentősége a molekuláris filogenetikának, amely a DNS-szekvencia alapján határozza meg két (vagy több) faj rokonságát. A növények tudományos besorolását a Nemzetközi Szabályzat az Algák, Gombák és Növények Nómenklatúrájára (International Code of Nomenclature for algae, fungi, and plants, ICN) határozza meg, amit a Nemzetközi Botanikai Kongresszus tart az új tudományos eredményeknek megfelelő színvonalon.[143][144]
Az aktuális rendszertani besorolás szerint a Növények (Plantae) országa az Eukarióták domén része. A további kategóriák: törzs (divisio), osztály (classis), rend (ordo), család (familia), nemzetség (genus), faj (species). Egy faj tudományos neve a nemzetség és a faj latin nevéből áll: pl. a fehér liliom (Lilium candidum) esetében a Lilium a nemzetség, a candidum a faj megnevezése.
A rokonság megállapítása sokszor nem egyszerű és túlmegy a felszínes hasonlóságokon. Például a Pereskia nemzetség a kaktuszokhoz tartozik, annak ellenére, hogy jól fejlett levelekkel rendelkező bokrok tartoznak hozzá; azonban a többi kaktuszhoz hasonlóan specializált areolákat és abból fejlődő tüskéket növeszt, bizonyítva az összetartozást.[145][146]

Pereskia aculeata

Echnocactus grusonii
Külső eltéréseik ellenére ez a két faj rokonságban áll egymással
A párhuzamos evolúció hasonló külalakot hozhat létre, például egyes kutyatejfélék a kaktuszokhoz hasonlóan vízraktározó, levéltelen szárakkal rendelkeznek, azonban virágaik alapján megbizonyosodhatunk róla, hogy nem állnak leszármazási rokonságban egymással.
Az 1990-es évek óta a taxonómiában a morfológiai jellegeket egyre inkább a DNS-szekvencia váltja fel vagyis a közvetett fenotípus helyett közvetlenül a genotípust, a genetikai kódot veszik figyelembe. Ennek köszönhető, hogy a gombákat, amelyeket korábban a növények egyik távoli alcsoportjaként soroltak be, ma inkább az állatvilághoz tartják közelebbinek.[147]
| |||||||||||||
1998-ban a Zárvatermő Filogenetikai Csoport az akkor hozzáférhető DNS-szekvenciák alapján közzétette a legtöbb virágos növény rokonsági kapcsolatait. Ennek eredményeképpen sok, addig vitatott kérdés rendeződött, például, hogy mely családok állnak közel a legkorábbi zárvatermőkhöz.[38] A rokonsági viszonyok tisztázása a növények evolúciós folyamatának megértését is segíti.[148] A munka azonban távolról sincs még befejezve, a molekuláris genetikai kutatások által szállított hatalmas mennyiségű adat és azok számítógépes feldolgozása tovább finomíthatja ismereteinket a növények rokonsági viszonyairól.[149]
Kapcsolódó cikkek
Jegyzetek
- ↑ Liddell & Scott 1940.
- ↑ Gordh & Headrick 2001, 134. o.
- ↑ Online Etymology Dictionary 2012.
- ↑ Bold 1977, 7. o.
- ↑ Judd et al. 2002, 2. o.
- ↑ Sumner 2000. 16. o.
- ↑ a b Reed 1942 7–29. o.
- ↑ Iyer 2009 117. o.
- ↑ Needham, Lu, Huang 1986
- ↑ a b Greene 1909 140–142. o.
- ↑ Bennett1 30. o.
- ↑ Mauseth 2003 532. o.
- ↑ Dallal 197. o.
- ↑ Panaino 93. o.
- ↑ Levey 116. o.
- ↑ Hill
- ↑ Yaniv & Bachrach 157. o.
- ↑ Sprague
- ↑ Waggoner
- ↑ Magyar füveskönyvek, füvészkönyvek
- ↑ Capon 220–223. o.
- ↑ Hoek, Mann & Jahns 9. o.
- ↑ Starr 299. o.
- ↑ Morton 377. o.
- ↑ Harris 76–81. o.
- ↑ de Candolle 2006, 9–25, 450–465. o.
- ↑ National Science Foundation 1989.
- ↑ Chaffey 2007, 481–482. o.
- ↑ Jasechko et al. 2013, 347–350. o.
- ↑ Nobel 1983, 608. o.
- ↑ Yates & Mather 1963, 91–129. o.
- ↑ Finney 1995, 554–573. o.
- ↑ Cocking 1993.
- ↑ Cousens & Mortimer 1995.
- ↑ Ehrhardt & Frommer 2012, 1–21. o.
- ↑ Haberlandt 1902.
- ↑ Leonelli et al. 2012.
- ↑ a b Burger 2013.
- ↑ Campbell et al. 2008, 186–187. o.
- ↑ Campbell et al. 2008, 1240. o.
- ↑ Gust 1996.
- ↑ Missouri Botanical Garden 2009.
- ↑ Ben-Menahem 5368. o.
- ↑ Chapman et al. 2001, 56. o.
- ↑ Braselton 2013.
- ↑ Campbell et al. 2008, 619–620. o.
- ↑ Capon 2005, 10–11. o.
- ↑ Mauseth 2003, 1–3. o.
- ↑ Acharya 440. o.
- ↑ Botanical Society of America 2013.
- ↑ Ben-Menahem 5367–5368. o.
- ↑ Butz 2007, 534–553. o.
- ↑ Zohary & Hopf 2000, 20–22. o.
- ↑ Floros, Newsome & Fisher 2010.
- ↑ Schoening 2005.
- ↑ Campbell et al. 2008, 190–193. o.
- ↑ Kim & Archibald 2009, 1–39. o.
- ↑ Howe et al. 2008, 2675–2685. o.
- ↑ Takaichi 2011, 1101–1118. o.
- ↑ a b Lewis & McCourt 2004, 1535–1556. o.
- ↑ Padmanabhan & Dinesh-Kumar 2010, 1368–1380. o.
- ↑ Schnurr et al. 2002, 1700–1709. o.
- ↑ Ferro et al. 2002, 11487–11492. o.
- ↑ Kolattukudy 1996, 83–108. o.
- ↑ Fry 1989, 1–11. o.
- ↑ Thompson & Fry 2001, 23–34. o.
- ↑ Kenrick & Crane 1997, 33–39. o.
- ↑ Benderoth 9118–9123. o.
- ↑ Jeffreys 2005, 38–40. o.
- ↑ University of Maryland Medical Center 2011.
- ↑ Klemm et al. 2005.
- ↑ Scharlemann & Laurance 2008, 52–53. o.
- ↑ Mauseth 2003, 786–818. o.
- ↑ Burrows 1990, 1–73. o.
- ↑ Addelson
- ↑ Grime & Hodgson 1987, 283–295. o.
- ↑ Mauseth 2003, 819–848. o.
- ↑ Herrera & Pellmyr 2002, 211–235. o.
- ↑ Proctor & Yeo 1973, 479. o.
- ↑ Herrera & Pellmyr 2002, 157–185. o.
- ↑ Herrera & Pellmyr 2002, 185–210. o.
- ↑ Bennett & Willis 2001, 5–32. o.
- ↑ Beerling 287–394. o.
- ↑ Björn et al. 1999, 449–454. o.
- ↑ Ben-Menahem 5369–5370. o.
- ↑ Ben-Menahem 5369. o.
- ↑ Stace 2010b, 629–633. o.
- ↑ Hancock 2004, 190–196. o.
- ↑ Sobotka, Sáková & Curn 2000, 103–112. o.
- ↑ Campbell et al. 2008, 495–496. o.
- ↑ Morgensen 1996, 383–384. o.
- ↑ Benderoth 9118–9123. o.
- ↑ Arabidopsis Genome Initiative 2000, 796–815. o.
- ↑ Devos & Gale 2000.
- ↑ University of California-Davis 2012.
- ↑ Russin et al. 1996, 645–658. o.
- ↑ Rochaix, Goldschmidt-Clermont & Merchant 1998, 550. o.
- ↑ Glynn et al. 2007.
- ↑ Possingham & Rose 1976, 295–305. o.
- ↑ Sun et al. 2002, 95–100. o.
- ↑ Heinhorst & Cannon 1993, 1–9. o.
- ↑ Schell & Van Montagu 1977, 159–179. o.
- ↑ Bird 2007, 396–398. o.
- ↑ Spector 2012, 8. o.
- ↑ Mauseth 2003, 552–581. o.
- ↑ Copeland 1938, 383–420. o.
- ↑ Woese et al. 1977, 305–311. o.
- ↑ Cavalier-Smith 2004, 1251–1262. o.
- ↑ Mauseth 2003, 617–654. o.
- ↑ Becker 999–1004. o.
- ↑ Fairon-Demaret 1996, 217–233. o.
- ↑ Stewart & Rothwell 1993, 279–294. o.
- ↑ Taylor, Taylor & Krings 2009, chapter 13.
- ↑ Mauseth 2003, 720–750. o.
- ↑ Mauseth 2003, 751–785. o.
- ↑ Lee et al. 2011, e1002411. o.
- ↑ Mauseth 2003, 278–279. o.
- ↑ Mauseth 2003, 280–314. o.
- ↑ Mauseth 2003, 315–340. o.
- ↑ Mauseth 2003, 341–372. o.
- ↑ Mauseth 2003, 373–398. o.
- ↑ Darwin 1880, 129–200. o.
- ↑ Darwin 1880, 449–492. o.
- ↑ Mauseth 2003, 411–412. o.
- ↑ Sussex 2008, 1189–1198. o.
- ↑ Campbell et al. 2008, 827–830. o.
- ↑ Mauseth 2003, 411–413. o.
- ↑ Taiz & Zeiger 2002, 461–492. o.
- ↑ Taiz & Zeiger 2002, 519–538. o.
- ↑ Lin, Zhong & Grierson 2009, 331–336. o.
- ↑ Taiz & Zeiger 2002, 539–558. o.
- ↑ Raven, Evert & Eichhorn 2005, 9. o.
- ↑ Mauseth 2003, 62–81. o.
- ↑ a b Campbell et al. 2008, 739. o.
- ↑ a b Campbell et al. 2008, 812–814. o.
- ↑ a b Mauseth 2003, 185–208. o.
- ↑ Mithila et al. 2003, 408–414. o.
- ↑ Campbell et al. 2008, 741. o.
- ↑ Mauseth 2003, 114–153. o.
- ↑ Mauseth 2003, 154–184. o.
- ↑ Mauseth 2003, 209–243. o.
- ↑ Lilburn et al. 2006.
- ↑ McNeill et al. 2011, Preamble, para. 7. o.
- ↑ Mauseth 2003, 528–551. o.
- ↑ Mauseth 2012, 446–449. o.
- ↑ Anderson 26–27. o.
- ↑ Mauseth 2012, 453. o.
- ↑ Chase et al. 2003, 399–436. o.
- ↑ Morton 1981, 459–459. o.
Irodalom
- ↑ Acharya: Acharya, Deepak, Shrivastava Anshu. Indigenous Herbal Medicines: Tribal Formulations and Traditional Herbal Practices (angol nyelven). Jaipur, India: Aavishkar Publishers (2008). ISBN 81-7910-252-1
- ↑ Addelson: Addelson, Barbara: Natural Science Institute in Botany and Ecology for Elementary Teachers (angol nyelven). Botanical Gardens Conservation International, 2003. December. [2013. május 23-i dátummal az eredetiből archiválva]. (Hozzáférés: 2013. június 8.)
- ↑ Anderson: Anderson, Edward F.. The Cactus Family (angol nyelven). Timber Press (2001). ISBN 978-0-88192-498-5
- ↑ Armstrong: Armstrong, J. E. Hearst (1996). „Carotenoids 2: Genetics and Molecular Biology of Carotenoid Pigment Biosynthesis” (angol nyelven). FASEB J. 10 (2), 228–37. o. PMID 8641556.
- ↑ Becker: Becker, Burkhard, Birger Marin (2009). „Streptophyte Algae and the Origin of Embryophytes”. Annals of Botany, Oxford 103 (7), 999–1004. o, Kiadó: Oxford University Press. DOI:10.1093/aob/mcp044. PMID 19273476. PMC 2707909. (Hozzáférés: 2013. június 16.)
- ↑ Beerling: Beerling, D. J., C. P. Osborne, W. G. Chaloner (2001). „Evolution of Leaf-form in Land Plants Linked to Atmospheric CO2 Decline in the Late Palaeozoic Era” (angol nyelven). Nature 410 (6826), 352–4. o. DOI:10.1038/35066546. PMID 11268207.
- ↑ Benderoth: Benderoth, Markus (2006. Június). „Positive Selection Driving Diversification in Plant Secondary Metabolism” (angol nyelven). Proceedings of the National Academy of Sciences of the United States of America, Washington D.C. 103 (24), 9118–23. o. DOI:10.1073/pnas.0601738103. PMID 16754868. JSTOR 30051907, PMC 1482576.
- ↑ Ben-Menahem: Ben-Menahem, Ari. Historical Encyclopedia of Natural and Mathematical Sciences (angol nyelven). Berlin: Springer-Verlag (2009). ISBN 3-540-68831-5
- ↑ Bennett1: Bennett, Charles E., William A. Hammond. The Characters of Theophrastus – Introduction (angol nyelven). London: Longmans, Green, and Co. (1902). Hozzáférés ideje: 2012. június 12.
- ↑ Bennett2: Bennett, K. D., K. J. Willis.szerk.: John P. Smol & H. John B. Birks: Pollen, Tracking Environmental Change Using Lake Sediments 3: Terrestrial, Algal, and Siliceous Indicators (angol nyelven). Dordrecht, Germany: Kluwer Academic Publishers (2001)
- Bird, Adrian (2007. május 1.). „Perceptions of Epigenetics”. Nature 447 (7143), 396–8. o. DOI:10.1038/nature05913. PMID 17522671.
- (1999. november 1.) „Ozone Depletion, Ultraviolet Radiation and Plant Life”. Chemosphere – Global Change Science, Philadelphia 1 (4), 449–454. o, Kiadó: Elsevier Ltd.. DOI:10.1016/S1465-9972(99)00038-0. (Hozzáférés: 2013. június 16.)
- Bold, H. C.. The Plant Kingdom, 4th, Englewood Cliffs, NJ: Prentice-Hall (1977). ISBN 0-13-680389-X
- Braselton, J. P.: What is Plant Biology?. Ohio University, 2013. [2015. szeptember 24-i dátummal az eredetiből archiválva]. (Hozzáférés: 2013. június 3.)
- Burger, William C.: Angiosperm Origins: A Monocots-First Scenario. The Field Museum, 2013. [2012. október 23-i dátummal az eredetiből archiválva]. (Hozzáférés: 2016. január 12.)
- Burrows, W. J.. Processes of Vegetation Change. London: Unwin Hyman (1990). ISBN 0-04-580013-8
- Butz, Stephen D.. Science of Earth Systems, 2, Clifton Park, NY: Delmar Cengage Learning (2007). ISBN 1-4180-4122-X
- Biology, 8, San Francisco: Pearson – Benjamin Cummings (2008). ISBN 978-0-321-54325-7
- de Candolle, Alphonse. Origin of Cultivated Plants. Glacier National Park, MT: Kessinger Publishing (2006). ISBN 978-1-4286-0946-4
- Capon, Brian. Botany for Gardeners, 2nd, Portland, OR: Timber Publishing (2005). ISBN 0-88192-655-8
- Cavalier-Smith, Thomas (2004). „Only Six Kingdoms of Life” (PDF). Proceedings of the Royal Society of London B 271 (1545), 1251–1262. o. DOI:10.1098/rspb.2004.2705. PMID 15306349.
- Chaffey, Nigel (2007). „Esau's Plant Anatomy, Meristems, Cells, and Tissues of the Plant Body: their Structure, Function, and Development”. Annals of Botany 99 (4), 785–786. o. DOI:10.1093/aob/mcm015.
- Science Web. Cheltenham, GB: Nelson Thornes (2001). ISBN 0-17-438746-6
- (2003) „An Update of the Angiosperm Phylogeny Group Classification for the Orders and Families of Flowering Plants: APG II” (PDF). Botanical Journal of the Linnean Society 141 (4), 399–436. o, Kiadó: The Linnean Society of London. DOI:10.1046/j.1095-8339.2003.t01-1-00158.x.
- (2007) „The JAZ Family of Repressors is the Missing Link in Jasmonate Signaling”. Nature 448 (7154), 666–71. o. DOI:10.1038/nature06006. PMID 17637675.
- Cocking, Edward C.. „Obituary: Professor F. C. Steward”, The Independent, 1993. október 18. (Hozzáférés: 2013. július 5.)
- Copeland, Herbert Faulkner (1938). „The Kingdoms of Organisms”. Quarterly Review of Biology 13 (4), 383–420. o. DOI:10.1086/394568.
- (2007. március 1.) „'Open Minded' Cells: How Cells Can Change Fate” (PDF)). Trends in Cell Biology 17 (3), 101–6. o. [2013. december 15-i dátummal az eredetiből archiválva]. DOI:10.1016/j.tcb.2006.12.005. PMID 17194589. (Hozzáférés: 2012. november 23.)
- Dynamics of Weed Populations. Cambridge: Cambridge University Press (1995). ISBN 978-0-521-49969-9
- Dallal, Ahmad. Islam, Science, and the Challenge of History. New Haven, CT: Yale University Press (2010). ISBN 978-0-300-15911-0
- Darwin, Charles. The Power of Movement in Plants (PDF), London: Murray (1880)
- (1962) „Isolement et détermination de la structure du jasmonate de méthyle, constituant odorant caractéristique de l'essence de jasminIsolement et détermination de la structure du jasmonate de méthyle, constituant odorant caractéristique de l'essence de jasmin”. Helvetica Chimica Acta 45 (2), 675–685. o. DOI:10.1002/hlca.19620450233.
- (2000. május 1.) „Genome Relationships: The Grass Model in Current Research”. The Plant Cell 12 (5), 637–646. o, Kiadó: American Society of Plant Physiologists. DOI:10.2307/3870991. PMID 10810140.
- (2012. február 1.) „New Technologies for 21st Century Plant Science” (PDF). The Plant Cell 24 (2), 374–94. o. DOI:10.1105/tpc.111.093302. PMID 22366161.
- Ereshefsky, Marc (1997). „The Evolution of the Linnaean Hierarchy”. Biology and Philosophy 12 (4), 493–519. o, Kiadó: Kluwer Academic Publishers. DOI:10.1023/A:1006556627052.
- (2002. augusztus 20.) „Integral Membrane Proteins of the Chloroplast Envelope: Identification and Subcellular Localization of New Transporters”. Proceedings of the National Academy of Sciences of the United States of America 99 (17), 11487–11492. o. DOI:10.1073/pnas.172390399.
- Fairon-Demaret, Muriel (1996. október 1.). „Dorinnotheca streelii Fairon-Demaret, gen. et sp. nov., a New Early Seed Plant From the Upper Famennian of Belgium”. Review of Palaeobotany and Palynology 93, 217–233. o. DOI:10.1016/0034-6667(95)00127-1.
- (1995. november 1.) „Frank Yates 12 May 1902 – 17 June 1994”. Biographical Memoirs of Fellows of the Royal Society 41, 554–573. o. DOI:10.1098/rsbm.1995.0033.
- Feeding the World Today and Tomorrow: The Importance of Food Science and Technology (PDF). Institute of Food Technologists, 2010. [2012. február 16-i dátummal az eredetiből archiválva]. (Hozzáférés: 2012. március 1.)
- Fry, S. C. (1989). „The Structure and Functions of Xyloglucan”. Journal of Experimental Biology, Cambridge 40, Kiadó: The Company of Biologists.
- A Dictionary of Entomology. Cambridge, MA: CABI Publishing (2001). ISBN 978-0-85199-291-4
- Scientific Papers of Asa Gray: Selected by Charles Sprague Sargent. Boston, MA: Houghton Mifflin (1889). Hozzáférés ideje: 2012. február 26.
- Greene, Edward Lee. Landmarks of botanical history: a study of certain epochs in the development of the science of botany: part 1, Prior to 1562 A.D.. Washington, D.C.: Smithsonian Institution (1909)
- (2007. május 1.) „Chloroplast Division”. Traffic 8 (5), 451–61. o. DOI:10.1111/j.1600-0854.2007.00545.x. PMID 17451550.
- (2010) „The Path from C3 to C4 Photosynthesis”. Plant Physiology 155 (1), 56–63. o. DOI:10.1104/pp.110.165308. PMID 20940348.
- (1987) „Botanical Contributions to Contemporary Ecological Theory”. The New Phytologist 106.
- Gust, Devens: Why Study Photosynthesis?. Arizona State University, 1996. [2012. február 9-i dátummal az eredetiből archiválva]. (Hozzáférés: 2012. február 26.)
- Hancock, James F.. Plant Evolution and the Origin of Crop Species. Cambridge, MA: CABI Publishing (2004). ISBN 0-85199-685-X
- Haberlandt, G. (1902). „Kulturversuche mit isolierten Pflanzenzellen” (german nyelven). Mathematisch-naturwissenschaftliche, Vienna 111 (1), 69–92. o, Kiadó: Akademie der Wissenschaften in Wien Sitzungsberichte.
- Harris, Henry. The Birth of the Cell. New Haven, CT: Yale University Press (2000). ISBN 0-300-08295-9
- (1993. január 1.) „DNA Replication in Chloroplasts”. Journal of Cell Science 104 (104), 1. o. (Hozzáférés: 2013. július 2.)
- Plant Animal Interactions: An Evolutionary Approach. Hoboken, NJ: Blackwell Science (2002). ISBN 978-0-632-05267-7
- Hill, Arthur W. (1915). „The History and Functions of Botanic Gardens”. Annals of the Missouri Botanical Garden 2 (1/2), 185–240. o. DOI:10.2307/2990033.
- Algae: An Introduction to Phycology. Cambridge: Cambridge University Press (2005). ISBN 0-521-30419-9
- (2008) „The Origin of Plastids”. Philosophical Transactions of the Royal Society B: Biological Sciences 363 (1504), 2675–85. o. DOI:10.1098/rstb.2008.0050. PMID 18468982.
- Hunter, Philip: What Genes Remember. Web.archive.org, 2008. május 1. [2008. május 1-i dátummal az eredetiből archiválva]. (Hozzáférés: 2013. augusztus 24.)
- Iyer, Meena. Faith & Philosophy of Zoroastrianism. Delhi, India: Kalpaz Publications (2009). ISBN 978-81-7835-724-9
- (2009. augusztus 4.) „A DNA Barcode for Land Plants”. Proceeding of the National Academy of Sciences (PNAS) 106 (31), 12794–7. o. DOI:10.1073/pnas.0905845106. PMID 19666622.
- (2013. április 3.) „Terrestrial Water Fluxes Dominated by Transpiration”. Nature 496 (7445), 347–50. o. DOI:10.1038/nature11983. PMID 23552893.
- Jeffreys, Diarmuid. Aspirin : The Remarkable Story of a Wonder Drug. New York: Bloomsbury (2005). ISBN 978-1-58234-600-7
- Plant Systematics, a Phylogenetic Approach. Sunderland, MA: Sinauer Associates (2002). ISBN 0-87893-403-0
- Karp, Gerald. Cell and Molecular Biology: Concepts and Experiments. Hoboken, NJ: John Wiley & Sons (2009). ISBN 978-0-470-48337-4
- (1997. szeptember 1.) „The Origin and Early Evolution of Plants on Land”. Nature 389 (6646), 33–39. o. DOI:10.1038/37918.
- Diversity and Evolution of Plastids and Their Genomes, The Chloroplast, Plant Cell Monographs. DOI: 10.1007/978-3-540-68696-5_1 (2009). ISBN 978-3-540-68692-7
- Klemm, Dieter (2005. szeptember 6.). „Cellulose: Fascinating Biopolymer and Sustainable Raw Material”. ChemInform, Hoboken, NJ 36 (36), Kiadó: John Wiley & Sons. DOI:10.1002/chin.200536238.
- Kolattukudy, Pappachan E.. 3, Plant Cuticles, Environmental Plant Biology Series. Oxford: BIOS Scientific Publishers Ltd (1996). ISBN 1-85996-130-4
- (2005. június 1.) „Use of DNA Barcodes to Identify Flowering Plants”. Proceeding of the National Academy of Sciences (PNAS) 102 (23), 8369–74. o. DOI:10.1073/pnas.0503123102. PMID 15928076. Supporting Information
- (2011) „A Functional Phylogenomic View of the Seed Plants”. PLOS Genetics 7 (12), e1002411. o. DOI:10.1371/journal.pgen.1002411. PMID 22194700.
- (2012) „Under One Leaf, A Historical Perspective on the UK Plant Science Federation”. New Phytologist 102 (23), 8369–74. o. DOI:10.1111/j.1469-8137.2012.4168.x. PMID 22530650.
- Lepp, Heino: Mosses. Australian National Botanic Gardens, 2012. (Hozzáférés: 2013. július 14.)
- Levey, Martin. Early Arabic Pharmacology: An Introduction Based on Ancient and Medieval Sources. Leiden: Brill Archive (1973). ISBN 978-90-04-03796-0
- (2004) „Green Algae and the Origin of Land Plants”. American Journal of Botany, St. Louis, MO 91 (10), 1535–56. o. DOI:10.3732/ajb.91.10.1535. PMID 21652308.
- Botane (βοτάνη). Oxford: Clarendon Press via Perseus Digital Library, Tufts University (1940)
- Lilburn, Timothy G. (2006). „Computational aspects of systematic biology”. Briefings in Bioinformatics 7 (2), 186–195. o. DOI:10.1093/bib/bbl005. PMID 16772262.
- (2009) „Recent Advances in Ethylene Research”. Journal of Experimental Botany, Oxford 60 (12), 3311–36. o. DOI:10.1093/jxb/erp204. PMID 19567479.
- (2003) „Phragmoplastin, Green Algae and the Evolution of Cytokinesis”. International Journal of Systematic and Evolutionary Microbiology, Reading, UK 53 (6), 1715–1718. o. DOI:10.1099/ijs.0.02561-0.
- (2002) „Evolution of Sucrose Synthesis”. Plant Physiology 128 (4), 1490–500. o. DOI:10.1104/pp.010898. PMID 11950997.
- Lüttge, Ulrich (2006). „Photosynthetic Flexibility and Ecophysiological Plasticity: Questions and Lessons from Clusia, the Only CAM Tree, in the Neotropics”. New Phytologist, Hoboken, NJ 171 (1), 7–25. o. DOI:10.1111/j.1469-8137.2006.01755.x. PMID 16771979.
- Mann, J.. Secondary Metabolism, 2nd ed.. Oxford: Oxford University Press (1987). ISBN 0-19-855529-6
- Mauseth, James D.. Botany : An Introduction to Plant Biology, 3rd, Sudbury, MA: Jones and Bartlett Learning (2003). ISBN 0-7637-2134-4
- Mauseth, James D.. Botany : An Introduction to Plant Biology, 5th, Sudbury, MA: Jones and Bartlett Learning (2012). ISBN 978-1-4496-6580-7
- International Code of Nomenclature for algae, fungi, and plants (Melbourne Code) adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011 [archivált változat]. A.R.G. Gantner Verlag KG (2011). ISBN 978-3-87429-425-6. Hozzáférés ideje: 2016. január 12. [archiválás ideje: 2013. november 4.]
- Medbury, Scot: Taxonomy and Arboreturm Design (PDF). Harvard University, 1993. [2015. június 9-i dátummal az eredetiből archiválva]. (Hozzáférés: 2013. július 26.)
- (2003. január 1.) „Thidiazuron Induces Shoot Organogenesis at Low Concentrations and Somatic Embryogenesis at High Concentrations on Leaf and Petiole Explants of African Violet (Saintpaulia ionantha Wendl)”. Plant Cell Reports 21 (5), 408–14. o. DOI:10.1007/s00299-002-0544-y. PMID 12789442.
- Morgensen, H. L. (1996). „The Hows and Whys of Cytoplasmic Inheritance in Seed Plants”. American Journal of Botany 83 (3). DOI:10.2307/2446172.
- Morton, Alan G.. History of Botanical Science: An Account of the Development of Botany from Ancient Times to the Present Day. London: Academic Press (1981). ISBN 978-0-12-508380-5
- Needham, Joseph. Science and Civilisation in China, Vol. 6 Part 1 Botany. Cambridge University Press (1986)
- Nobel, P. S.. Biophysical Plant Physiology and Ecology. San Francisco: W. H. Freeman (1983). ISBN 0-7167-1447-7
- Oberlies, Thomas. Die Religion des Rgveda (german nyelven). Sammlung De Nobili (1998). ISBN 978-3-900271-31-2
- (2010) „All Hands on Deck—The Role of Chloroplasts, Endoplasmic Reticulum, and the Nucleus in Driving Plant Innate Immunity”. Molecular Plant-Microbe Interactions, St. Paul, MN 23 (11), 1368–80. o, Kiadó: The American Phytopathological Society. DOI:10.1094/MPMI-05-10-0113. PMID 20923348.
- Panaino, Antonio. Ideologies as Intercultural Phenomena: Proceedings of the Third Annual Symposium of the Assyrian and Babylonian Intellectual Heritage Project, Held in Chicago, USA, October 27–31, 2000. Bologna: Mimesis Edizioni (2002). ISBN 978-88-8483-107-1
- Mosses and Liverworts. London: HarperCollins UK (2005). ISBN 978-0-00-220212-1
- (1976. május 18.) „Chloroplast Replication and Chloroplast DNA Synthesis in Spinach Leaves” (PDF). Proceedings of the Royal Society B: Biological Sciences 193 (1112), 295–305. o. DOI:10.1098/rspb.1976.0047.
- The Pollination of Flowers, New Naturalist series. London: Harper Collins (1973). ISBN 0-00-219504-6
- Biology of Plants, 7th, New York: W. H. Freeman (2005). ISBN 0-7167-1007-2
- Reed, Howard S.. A Short History of the Plant Sciences. New York: Ronald Press (1942)
- Reik, Wolf (2007. május 1.). „Stability and Flexibility of Epigenetic Gene Regulation in Mammalian Development”. Nature 447 (7143), 425–32. o. DOI:10.1038/nature05918. PMID 17522676.
- (1995) „Dioecy and its Correlates in the Flowering Plants”. American Journal of Botany 82 (5), 596. o. DOI:10.2307/2445418.
- The Molecular Biology of Chloroplasts and Mitochondria in Chlamydomonas. Dordrecht, Germany: Kluwer Academic (1998). ISBN 978-0-7923-5174-0
- Roux, Stanley J. (1984). „Ca2+ and Phytochrome Action in Plants”. BioScience, Berkeley, CA 34 (1), 25–9. o. DOI:10.2307/1309422. PMID 11540810.
- (1996) „Modification of a Specific Class of Plasmodesmata and Loss of Sucrose Export Ability in the sucrose export defective1 Maize Mutant” (PDF). The Plant Cell 8 (4), 645–658. o. DOI:10.1105/tpc.8.4.645. PMID 12239395.
- Sattler, R. (1992). „Process morphology: structural dynamics in development and evolution” (PDF). Canadian Journal of Botany 70 (4), 708–714. o. DOI:10.1139/b92-091.
- (1992) „Multivariate analysis confirms the continuum view of plant form”. Annals of Botany 69, 249–262. o.
- Savidan, Y. H. (2000). „Apomixis: Genetics and Breeding”. Plant Breeding Reviews 18, 13–86. o. DOI:10.1002/9780470650158.ch2.
- Scharf, Sara T. (2009). „Identification Keys, the "Natural Method," and the Development of Plant Identification Manuals”. Journal of the History of Biology 42 (1), 73–117. o. DOI:10.1007/s10739-008-9161-0. PMID 19831202.
- (2008) „How Green are Biofuels?”. Science 319 (5859), 43–4. o, Kiadó: American Association for the Advancement of Science. DOI:10.1126/science.1153103. PMID 18174426.
- (1977) „The Ti-plasmid of Agrobacterium tumefaciens, a Natural Vector for the Introduction of Nif Genes in Plants?”. Basic Life Sciences 9, 159–79. o. DOI:10.1007/978-1-4684-0880-5_12. PMID 336023.
- Schoening, Steve: California Noxious and Invasive Weed Action Plan (PDF). California Department of Food and Agriculture, 2005. (Hozzáférés: 2012. március 1.)
- (2002) „Fatty Acid Export from the Chloroplast. Molecular Characterization of a Major Plastidial Acyl-Coenzyme a Synthetase from Arabidopsis”. Plant Physiology 129 (4), 1700–9. o. DOI:10.1104/pp.003251. PMID 12177483.
- Silyn-Roberts, Heather. Writing for Science and Engineering: Papers, Presentation. Oxford: Butterworth-Heinemann (2000). ISBN 0-7506-4636-5
- Small, Michael. Dynamics of Biological Systems. Boca Raton, FL: CRC Press (2012). ISBN 978-1-4398-5336-8
- (2000) „Molecular Mechanisms of Self-incompatibility in Brassica”. Current Issues in Molecular Biology 2 (4), 103–12. o. PMID 11471754.
- Spector, Tim. Identically Different: Why You Can Change Your Genes. London: Weidenfeld & Nicolson (2012). ISBN 978-0-297-86631-2
- Sprague, T. A. (1939). „The Herbal of Valerius Cordus”. The Journal of the Linnean Society of London LII (341), 1–113. o, Kiadó: Linnean Society of London. DOI:10.1111/j.1095-8339.1939.tb01598.x.
- Stace, Clive A. (2010a). „Classification by molecules: What's in it for field botanists?”. Watsonia 28. [2011. július 26-i dátummal az eredetiből archiválva]. (Hozzáférés: 2013. július 6.)
- Stace, Clive. New Flora of the British Isles, 3rd, Cambridge: Cambridge University Press (2010b). ISBN 978-0-521-70772-5
- Starr, Cecie. The Unity and Diversity of Life, AP, Belmomt, CA: Brooks/Cole, Cenpage Learning (2009). ISBN 978-1-111-58097-1 [halott link]
- Paleobiology and the Evolution of Plants. Cambridge: Cambridge University Press (1993). ISBN 978-0-521-38294-6
- Bananas, 3rd, Harlow, England: Longman (1987). ISBN 978-0-582-46357-8
- Sumner, Judith. The Natural History of Medicinal Plants. New York: Timber Press (2000). ISBN 0-88192-483-0
- (2002) „Crystal Structure of Pea Toc34, a Novel GTPase of the Chloroplast Protein Translocon”. Nature Structural Biology 9 (2), 95–100. o. DOI:10.1038/nsb744. PMID 11753431.
- Sussex, I. (2008). „The Scientific Roots of Modern Plant Biotechnology” (PDF). The Plant Cell 20 (5), 1189–98. o. DOI:10.1105/tpc.108.058735. PMID 18515500.
- Plant Physiology, 3rd, Sunderland, MA: Sinauer Associates (2002). ISBN 0-87893-823-0
- Takaichi, Shinichi (2011. június 1.). „Carotenoids in Algae: Distributions, Biosyntheses and Functions”. Marine Drugs 9 (12), 1101–1118. o. DOI:10.3390/md9061101. PMID 21747749.
- (1935) „The Use and Abuse of Vegetational Terms and Concepts”. Ecology, Washington, D.C. 16 (3), 284. o, Kiadó: Ecological Society of America. DOI:10.2307/1930070.
- Taylor, T.N.. Paleobotany, The Biology and Evolution of Fossil Plants, 2nd, Academic Press (2009). ISBN 978-0-12-373972-8
- (2001) „Restructuring of Wall-bound Xyloglucan by Transglycosylation in Living Plant Cells”. The Plant Journal, West Sussex, England 26 (1), 23–34. o, Kiadó: John Wiley & Sons. DOI:10.1046/j.1365-313x.2001.01005.x. PMID 11359607.
- Waggoner, Ben: University of California Museum of Paleontology. University of California-Berkeley, 2001. (Hozzáférés: 2012. február 27.)
- Phytohormones (PDF), New York: Macmillan (1937)
- (1997) „The Ecosystem: An Evolving Concept Viewed Historically”. Functional Ecology, London 11 (2), 268–271. o, Kiadó: British Ecological Society. DOI:10.1111/j.1365-2435.1997.00081.x.
- (1977. augusztus 1.) „An Ancient Divergence Among the Bacteria”. Journal of Molecular Evolution 9 (4), 305–311. o. DOI:10.1007/BF01796092. PMID 408502.
- Handbook of Medicinal Plants. Binghampton, NY: Haworth Press (2005). ISBN 1-56022-994-2
- Ronald Aylmer Fisher 1890–1962 (1963)
- Domestication of Plants in the Old World, 3rd, Oxford: Oxford University Press (2000). ISBN 978-0-19-850356-9
- The Arabidopsis Genome Initiative (2000). „Analysis of the Genome Sequence of the Flowering Plant Arabidopsis thaliana”. Nature, London 408 (6814), 796–815. o, Kiadó: Nature Publishing Group. DOI:10.1038/35048692. PMID 11130711.
- Auxins. Plant Hormones, Long Ashton Research Station, Biotechnology and Biological Sciences Research Council. [2021. december 10-i dátummal az eredetiből archiválva]. (Hozzáférés: 2013. július 15.)
- A Basic Introduction to the Science Underlying NCBI Resources. National Center for Biotechnology Information, 2004. március 30. (Hozzáférés: 2012. március 5.)
- Botany. Online Etymology Dictionary, 2012. (Hozzáférés: 2012. február 24.)
- Early Herbals – The German Fathers of Botany. National Museum of Wales, 2007. július 4. [2012. június 29-i dátummal az eredetiből archiválva]. (Hozzáférés: 2012. február 19.)
- Katherine Esau. National Science Foundation, 1989. (Hozzáférés: 2013. június 26.)
- Evolution and Diversity, Botany for the Next Millennium: I. The Intellectual: Evolution, Development, Ecosystems. Botanical Society of America. (Hozzáférés: 2013. június 25.)
- Herbal Medicine. University of Maryland Medical Center. [2012. március 2-i dátummal az eredetiből archiválva]. (Hozzáférés: 2012. március 2.)
- Paleobotany. Cleveland Museum of Natural History. (Hozzáférés: 2014. július 30.)
- Physical Map of Brachypodium'. University of California-Davis. [2016. március 5-i dátummal az eredetiből archiválva]. (Hozzáférés: 2012. február 26.)
- Plants and Life on Earth. Missouri Botanical Garden, 2009. (Hozzáférés: 2012. március 10.)
Fordítás
- Ez a szócikk részben vagy egészben a Botany című angol Wikipédia-szócikk ezen változatának fordításán alapul. Az eredeti cikk szerkesztőit annak laptörténete sorolja fel. Ez a jelzés csupán a megfogalmazás eredetét és a szerzői jogokat jelzi, nem szolgál a cikkben szereplő információk forrásmegjelöléseként.
 Biológiaportál • összefoglaló, színes tartalomajánló lap
Biológiaportál • összefoglaló, színes tartalomajánló lap








