Fosforilazione ossidativa
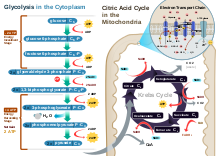
La fosforilazione ossidativa è un processo biochimico cellulare fondamentale e ubiquitario per la produzione di energia tramite la sintesi di ATP. Si tratta della fase finale della respirazione cellulare, dopo glicolisi, decarbossilazione ossidativa del piruvato e ciclo di Krebs.
L'ubicazione fisica del processo è sempre a cavallo di una membrana: negli eucarioti avviene presso la membrana interna mitocondriale, nei procarioti presso la membrana cellulare.
È composta da due fasi:
- Catena di trasporto degli elettroni: in questo processo gli elettroni trasportati da NADH e FADH2 vengono scambiati dalla catena enzimatica transmembrana generando un gradiente protonico
- sintesi di ATP tramite fosforilazione di ADP da parte dell'enzima ATP sintasi
Catena di trasporto degli elettroni
I coenzimi "caricati" nella matrice mitocondriale durante il ciclo di Krebs e nel citoplasma durante la glicolisi sono utilizzati nella cosiddetta catena di trasporto degli elettroni. Essa "scarica" i due coenzimi NADH e FADH2 (ossidandoli a NAD+ e FAD), trasferendone gli elettroni ad alcune molecole di trasporto, essenzialmente citocromi (particolari complessi proteici enzima-coenzima in grado di legare l'ossigeno). I citocromi sono deputati al trasporto degli elettroni verso gli enzimi NADH-Q reduttasi, succinato deidrogenasi, citocromo reduttasi e citocromo c ossidasi, capaci di "pompare" gli ioni H+ derivanti dall'ossidazione di NAD e FAD dalla matrice allo spazio intermembrana tramite un trasporto attivo attraverso la membrana interna, generando un gradiente protonico. I protoni non possono rientrare nella matrice per ristabilire l'equilibrio perché la membrana interna non è spontaneamente permeabile al loro passaggio. Il rientro avviene alla fine del processo per mezzo dell'enzima ATP-sintasi, che sfrutta l'energia del loro potenziale elettrochimico per la sintesi di ATP (vedi sotto). Dalla riduzione dell'accettore finale degli elettroni, l'ossigeno, e dagli ioni H+ derivano molecole di acqua che si aggiungono a quelle prodotte dalla glicolisi.
Generazione di ATP
Quando gli elettroni arrivano all'ossigeno, la catena di trasporto si interrompe per una frazione di tempo necessaria affinché avvenga il riequilibrio protonico. Si annulla la differenza di potenziale e di pH e i protoni ritornano nella matrice attraverso il complesso proteico (proteina canale) dell'ATP sintasi (chemiosmosi). L'energia potenziale liberata attiva la reazione endoergonica tra ADP e gruppi fosfato con sintesi di ATP. La reazione è:
ADP3- + H+ + Pi ATP4- + H2O.

Si avrà quindi l'ultima forma di conservazione di energia, immagazzinata nei legami di 38 molecole di ATP attraverso glicolisi, ciclo di Krebs e fosforilazione ossidativa. Va ricordato che questo è il rendimento massimo teorico di una molecola di glucosio, ma quello reale si aggira fra 36 e 32 molecole di ATP, per una minima permeabilità delle membrane agli elettroni e per la minor resa reale di NADH e FADH2, che è rispettivamente di 2,5 e 1,5 ATP e non 3 e 2 (v. sotto "Stechiometria"). La cellula potrà liberare al bisogno l'energia contenuta nell'ATP tramite il processo dell'idrolisi, catalizzato dall'enzima ATPasi Mg++ dipendente.
Dopo la sintesi di ATP per ristabilire la concentrazione di NADH all'interno della matrice si sfrutta il sistema navetta malato-aspartato, un sistema di due trasportatori. Il NADH non può attraversare la membrana mitocondriale quindi attraverso questa vengono trasportati gli elettroni provenienti dal NADH. Nel citosol il NADH si ossida e trasforma l'ossalacetato in malato grazie all'enzima malato deidrogenasi. Quest'ultimo attraversa la membrana mitocondriale interna e viene riossidato dal NAD+. L'ossalacetato tramite una reazione di transamminazione con il glutammato diventa aspartato grazie all'enzima aspartato transaminasi e viene trasportato nello spazio intermembrana.
Stechiometria
Per ogni molecola di NADH ossidata vengono prodotti in totale 2,5 molecole di ATP. Per ogni molecola di FADH2 sono invece prodotte 1,5 molecole di ATP.
Attraverso l'ossidazione glicolitica di una molecola di glucosio, la successiva conversione delle molecole di piruvato in acetil-CoA e infine il ciclo di Krebs sono sintetizzate in tutto 10 molecole di NADH e 2 di FADH2. Il valore energetico immagazzinato in queste molecole viene convertito in un totale di 28 molecole di ATP [(10x2,5) + (2x1,5)].
È importante però considerare che questi valori sono soltanto teorici, mentre in realtà la resa della reazione di sintesi di ATP è inferiore, per via della permeabilità, seppure bassa, della membrana ai protoni.
Inibitori
Alcune molecole sono in grado di inibire alcuni passaggi della fosforilazione ossidativa. Per questo motivo, esse sono a tutti gli effetti delle tossine.
- Il cianuro blocca la catena di trasporto degli elettroni perché si lega al Fe3+ del citocromo a3 più efficientemente dell'ossigeno, impedendo al citocromo di combinare gli elettroni con l'ossigeno.
- L'oligomicina inibisce la ATP sintasi, impedendole di ricaricare molecole ATP attraverso il gradiente protonico.
- La CCCP (m-cloro-carbonilcianuro-fenilidrazina) distrugge il gradiente protonico permettendo ai protoni di attraversare la membrana. Senza il gradiente, infatti, la ATP sintasi non è in grado di funzionare.
- Anche un detergente (o una sostanza in grado di distruggere le membrane cellulari) è in grado di distruggere il gradiente protonico, semplicemente eliminando la membrana a cavallo del quale si genera.
- Il rotenone impedisce il trasferimento di elettroni dai centri Fe-S del complesso I all'ubichinone. Gli elettroni che fanno il loro ingresso nella catena attraverso tale complesso sono quelli provenienti dal NADH e costituiscono la parte preponderante del potenziale in grado di generare gradiente protonico.
Per ognuna di queste tossine, l'azione tossica non si confina solo al processo di fosforilazione ossidativa, ma ha effetti a catena su gran parte del metabolismo cellulare. Se si somministra oligomicina, ad esempio, gli H+ non sono in grado di passare attraverso la ATP sintasi. Ciò induce una stabilizzazione del gradiente protonico, che alla lunga rende inefficaci le pompe protoniche, che non funzionano in presenza di gradienti eccessivamente elevati. Se non funzionano tali pompe, le molecole di NADH e FADH2 non saranno più nuovamente ossidate a NAD+ e FAD. Ciò significa che gli altri processi metabolici che se ne servono, come il ciclo di Krebs e la glicolisi, non saranno più in grado di funzionare.
Composti reattivi dell'ossigeno
Durante la fosforilazione ossidativa si possono formare composti transienti dell'ossigeno con una forte reattività, ovvero perossido di idrogeno, l'anione superossido e il radicale idrossile. La citocromo c ossidasi, complesso IV, è però in grado di mantenere il controllo sterico sulle molecole create, riducendo l'ossigeno ad acqua.
Peter Dennis Mitchell
«L'aspetto dell'attuale situazione di consenso che ho trovato più rimarchevole ed ammirevole è l'altruismo e la generosità con cui i primi oppositori dell'ipotesi chemiosmotica non solo l'hanno accettata, ma hanno attivamente contribuito a promuoverla allo status di teoria.»
(Peter Mitchell, Discorso alla cerimonia del premio Nobel, 1978)
La fosforilazione ossidativa è rimasta un mistero fino agli inizi degli anni 60 ed è grazie al biochimico inglese Peter D. Mitchell (e a successivi ricercatori) se ora conosciamo nel dettaglio questo processo. Grazie a questa ricerca fu insignito del Premio Nobel per la chimica nel 1978.
Bibliografia
- Lehninger, D., Cox, M. Principles of Biochemistry 3rd Ed., Worth Publications, New York, NT., 2001.
- David L. Nelson, Michael M. Cox, I Principi di Biochimica di Lehninger, 3ª ed., Bologna, Zanichelli, febbraio 2002, ISBN 88-08-09035-3.
- Jeremy M. Berg, John L. Tymoczko; Lubert Stryer, Biochimica, 5ª ed., Bologna, Zanichelli, ottobre 2003, ISBN 88-08-07893-0.
Voci correlate
- Respirazione cellulare
- Catena di trasporto degli elettroni
- Sistema shuttle del malato-aspartato
- Sistema shuttle del glicerolo fosfato
- Gradiente protonico elettrochimico transmembrana
- Proteina disaccoppiante
Altri progetti
Altri progetti
- Wikimedia Commons
 Wikimedia Commons contiene immagini o altri file su Fosforilazione ossidativa
Wikimedia Commons contiene immagini o altri file su Fosforilazione ossidativa
Collegamenti esterni
- (EN) Modello molecolare interattivo della succinato deidrogenasi, su ufp.pt. URL consultato l'8 agosto 2006 (archiviato dall'url originale il 25 settembre 2006).
- (EN) Modello molecolare interattivo del coenzima Q - citocromo c riduttasi, su ufp.pt. URL consultato l'8 agosto 2006 (archiviato dall'url originale il 5 agosto 2008).
- (EN) Modello molecolare interattivo della citocromo c ossidasi, su ufp.pt. URL consultato l'8 agosto 2006 (archiviato dall'url originale il 25 settembre 2006).
V · D · M | |
|---|---|
| Metabolismo | Via metabolica · Catena metabolica · Ciclo metabolico · Via anfibolica |
| Catabolismo | Glicolisi · Beta ossidazione · Decarbossilazione ossidativa del piruvato · Ciclo di Krebs · Fosforilazione ossidativa · Glicogenolisi · Chetogenesi |
| Anabolismo | Gluconeogenesi · Sintesi proteica · Sintesi degli acidi grassi · Glicogenosintesi · Via dei pentoso fosfati |
| Altre voci | Ciclo di Cori · Degradazione degli amminoacidi · Fotosintesi clorofilliana · Fotorespirazione · Ciclo di Calvin · Ciclo dell'urea · Fermentazione · Ciclo del gliossilato |
 Portale Biologia
Portale Biologia Portale Chimica
Portale Chimica Portale Medicina
Portale Medicina










